Проблемі лікування посттравматичної кісткової дисрегенерації присвячено значну кількість робіт, у яких розглядаються як хірургічні, так і нехірургічні методи стимуляції репаративного остеогенезу. Однак, незважаючи на досить високий відсоток успішного лікування хворих із порушенням процесів консолідації кісткових уламків, у 4,8 – 15,4% хворих досягти позитивних результатів не вдається. Найбільші труднощі при лікуванні порушень остеорепарації виникають після травм, отриманих внаслідок впливу високоенергетичного агента, що травмує. За даними різних джерел, у 10,8 – 13,7% хворих з високоенергетичними травмами нижніх кінцівок переломи не зросталися протягом 1,5 – 3 років. Відомо, що формування кісткової мозолі в області перелому відбувається завдяки клітинам-попередникам періосту, ендосту та мезенхімальним стромальним (стволовим) клітинам кістковомозкової порожнини (МСК КП). Одним з визначальних механізмів формування кісткової мозолі є остеогенне диференціювання клітин-попередників, що проліферують у кісткову рану. Дослідження останніх років показали, що диференціація МСК за остеобластичним типом є складним, багатогранним процесом і здійснюється під контролем факторів транскрипції, клітинних і матриксних взаємодій, системних і місцевих факторів. На процеси остеогенної диференціації значний вплив дуже багато місцевих факторів, переважна більшість яких продукується клітинами остеобластической лінії. Але, незважаючи на те, що в процесі остеогенного диференціювання клітин-попередників, що мігрують у кісткову рану, провідне значення відіграють паракринні фактори, що продукуються вже мають остеогенну спрямованість клітинами периоста і ендоста, робіт, присвячених вивченню змін морфофункціонального стану клітинних джерел остеорепарації в залежності від механізму травм знайти не вдалося.
У зв'язку з цим було проведено вивчення ролі змін морфофункціонального стану клітинних джерел остеорепарації (періосту, ендосту та мезенхімальних стромальних клітин кістково-мозкової порожнини (МСК КП) у порушенні репаративного остеогенезу. Для цього були проведені експериментальні та клінічні дослідження. Вивчали морфологічну картину. остеорепарації (періоста, ендосту та МСК КП).
Експериментальні дослідження проводилися in vivo та in vitro на 80 статевозрілих білих безпородних щурах самцях. Тварин підрозділили на три групи – контрольну, першу та другу. Контрольну групу утворили десять тварин. У цих тварин за допомогою МТТ-аналізу вивчали проліферативну активність клітин періоста, ендосту та МСК КП. Для чого після виведення з експерименту у інтактних тварин в асептичних умовах здійснювали відчленування обох гомілок і виділення з м'яких тканин гомілкових кісток. Берцеві кістки поміщали у спеціальний консервант та витримували в ньому протягом доби. Потім у стерильних умовах ножицями відсікали метаепіфізарні області, залишаючи для подальшого дослідження лише діафізарну частину кістки. Для визначення проліферативної активності МСК КП у стерильних умовах з кістковомозкового каналу за допомогою інсулінового шприца та фізіологічного розчину вимивали кістковомозковий вміст у чашки Петрі з культуральним середовищем. Для спрощення цієї процедури діафізарну частину кістки розтинали поперечно навпіл. Половина кісткових фрагментів розсікалася поздовжньо очними ножицями, ретельно відмивали від залишків кістковомозкового вмісту фізіологічним розчином і поміщали в чашки Петрі з колагеновою підкладкою і живильним середовищем таким чином, щоб з колагеновою підкладкою контактувала тільки окістя (мал. Таке становище кісткових фрагментів у чашках Петрі створювало умови для проліферації лише клітин окістя. Половину кісткових фрагментів, що залишилася, за допомогою скальпеля, ретельно очищали від окістя, промивали фізіологічним розчином і поміщали в чашки Петрі з колагеновою підкладкою і живильним середовищем таким чином, щоб з колагеновою підкладкою контактувала кісткова тканина (рис. 1 б). Це створювало умови для проліферації лише клітин власне кісткової тканини (клітин ендосту).

У тварин першої (35 щурів) та другої (35 щурів) груп моделювали низько- та високоенергетичні переломи гомілкової кістки. При моделюванні низькоенергетичних травм (перша група) в асептичних умовах під ефірним наркозом в області середньої третини гомілки скальпелем здійснювали розріз шкіри і виділяли гомілкову кістку. Вузьким долотом виконували остеотомію кістки в середній третині, після чого зашивали рану наглухо. При моделюванні високоенергетичних переломів (2 група) під ефірним наркозом області середньої третини гомілки за допомогою пружинного пристрою завдавали дозований удар, сила якого становила в середньому 2,55±0,04 кг×м/с. У 10 тварин (по 5 тварин кожної групи) травмовані сегменти фіксували параосально з використанням голок для внутрішньом'язових ін'єкцій та поліхлорвінілових трубок для внутрішньовенних інфузій. Виведення цих тварин з експерименту здійснювали на 7 добу після травми. Після виведення з експерименту у тварин в асептичних умовах відчленовували травмований сегмент, звільняли його від шкіри, фіксували в 10% розчині нейтрального формаліну, декальцинували і фарбували гематоксиліном та еозином. Отримані гістологічні препарати вивчали у світловому мікроскопі. Шістдесят тварин (по 30 тварин кожної групи) було виведено з експерименту через 1 годину після травми. Після виведення з експерименту у тварин в асептичних умовах відчленовували травмований сегмент, кісткові фрагменти акуратно виділяли з м'яких тканин та поміщали у флакон зі стерильним консервантом. Кісткові фрагменти витримували у флаконі з консервантом протягом однієї доби. Потім з кістковими фрагментами кожної з тварин виконували ті ж маніпуляції, що і у тварин контрольної групи і культивували в 22-інкубаторі за стандартних умов. На 5 – 8-му добу культивування за допомогою фазово-контрастної мікроскопії здійснювали підрахунок проліферуючих від кісткових фрагментів клітин періоста та ендосту. Клітини підраховували у зору мікроскопа при 100-кратном збільшенні на п'яти рівнях від лінії перелому. Як критерій, що визначає рівень підрахунку, використовували поле зору мікроскопа. На 14-ту добу культивування в 45 чашках Петрі кожної групи (по 15 чашок для кожної з клітинних культур) методом МТТ-аналізу визначали проліферативну активність клітин періоста, ендосту та МСК КП. В решті 45-ти чашок Петрі кожної групи (по 15 чашок для кожної з клітинних культур), після формування моношару за допомогою фазово-контрастної мікроскопії оцінювали морфологічний стан проліферуючих клітин окістя, ендосту та МСК КП. У моношарі культур періоста та ендосту за допомогою реакції з BCIP/NBT Liquid Substrate System (Sigma, США) визначали наявність або відсутність продукції клітинами лужної фосфатази (ЩФ), що дозволяло судити про наявність або відсутність остеогенної детермінованості (спрямованості) клітин.
Клінічні дослідження були проведені у 5-ти хворих із тривалим незрощенням переломів довгих кісток нижніх кінцівок. У 2-х хворих були незрощені переломи стегнової кістки, у 3-х хворих - великогомілкової кістки. Тривалість порушення цілісності кісткової тканини у хворих була від 11 місяців до 2 років. Під час оперативного лікування у хворих із проксимального кісткового фрагмента в області перелому здійснювали забір періостальних тканин розміром 0,5'0,5 см. Біопсійний матеріал поміщали у флакон із консервантом та витримували протягом доби. Потім у стерильних умовах ділянки периоста витягали з консерванту і поміщали в чашки Петрі з живильним середовищем і культивували в <2>2-інкубаторі за стандартних умов. Після появи клітинної проліферації періостальні тканини витягувалися, а клітини, що прикріпилися до пластику, культивувалися до формування моношару. За допомогою фазово-контрастної мікроскопії оцінювали морфологічний стан проліферуючих клітин періосту, а за допомогою реакції на ЛФ – їхню остеогенну детермінованість.
Фотодокументування клітинних культур та гістопрепаратів здійснювали за допомогою інвертованого мікроскопа Leica DMIL, робочої станції з обробки зображень Leica QWin500 Standart (версія 2.3, сер. № 3069) та відеокамери Sanyo TK-C1380 (Німеччина). Отриманий у результаті досліджень цифровий матеріал піддавали статистичній обробці.
При вивченні гістоморфологічних препаратів області кісткової рани на 7-му добу після травми було зазначено, що стан кісткових фрагментів та зрощення переломів у тварин першої та другої груп було по-різному. У тварин першої групи кісткові фрагменти мали значну кількість клітинних елементів із добре фарбованими ядрами. В області кісткової рани відбувалося утворення примітивних кісткових балок, характерних для формування кісткової мозолі (рис. 2а). У другій групі кількість клітинних елементів у кістковій тканині уламків було набагато менше, ніж у першій групі. Вони розташовувалися переважно у віддалених від лінії перелому ділянках кісткової тканини. У кістки, що прилягає безпосередньо до лінії перелому, зустрічалися поодинокі клітини із слабо профарбованими ядрами. Ці зміни морфологічно нагадували картину остеонекрозу. В області перелому репаративні процеси виявлялися у вигляді формування грубоволокнистої ретикулярної сполучної тканини, що заповнює кісткову рану (див. рис. 2б).

Таким чином, дані гістоморфологічних досліджень свідчать про те, що інтенсивність впливу на кістку травмуючого агента впливає стан кісткової тканини і протягом зрощення перелому. При низькоенергетичних травмах у кістковій рані репаративні процеси протікають із формуванням специфічного кісткового регенерату, а при високоенергетичних травмах репаративні процеси обмежуються утворенням неспецифічного рубцевого регенерату.
При культивуванні клітин окістя активна клітинна проліферація починалася на 5-6 добу. У тварин першої групи клітини окістя проліферували рівномірно вздовж всього кісткового фрагмента (рис 3а). У тварин другої групи проліферація клітин окістя вздовж кісткового фрагмента була нерівномірною. У зоні, що прилягає до лінії перелому, проліферували поодинокі клітини. Активна клітинна проліферація відзначалася у зоні, найбільш віддаленій від лінії перелому (рис. 3б).

Результати підрахунку кількості клітин періосту у тварин 1 та 2 груп представлені у таблиці 1 та на діаграмі (рис. 4).

Наведені дані свідчать про те, що у тварин першої групи кількість проліферуючих клітин була однаковою по всьому фрагменту кістки і становила 27-29 клітин у полі зору. У тварин другої групи кількість проліферуючих клітин у зоні перелому не перевищувала 6 у полі зору. У більш віддалених від лінії зламу зонах кількість проліферуючих клітин поступово зростала від 17 до 54 клітин. Відмінності в кількості клітин між рівнями від лінії перелому у тварин другої групи були вірогідними (між рівнями 1, 2 і 3 - t=2,39, p<0,05; між рівнями 3 і 4 - t=2,3, p<0,05; між рівнями 4 і 5 - t=2,2, p<0,05).
Зіставлення кількості клітин на 1 – 5-му рівнях у тварин двох груп показало, що у тварин першої групи, на перших трьох рівнях кількість клітин, що проліферують, значно перевищувала таку у тварин другої групи (р< 0,002 і р < 0,05). На четвертому рівні кількість проліферуючих клітин окістя в обох групах була приблизно однаковою (р> 0,10), а на п'ятому рівні від лінії перелому відмінності мали зворотну залежність. У цій галузі кількість проліферуючих клітин у другій групі була достовірно більшою, ніж у першій групі (р<0,05).
У процесі культивування проліферативна активність клітин періосту у тварин двох груп була вищою за контрольне значення. Проліферативна активність клітин періоста в першій групі була в 1,6 рази (t= 7,8; р< 0,001), а в другій в 1,4 раза (t= 7,7; р< 0,001) більше контрольних значень. Відмінності між проліферативною активністю клітин періоста першої та другої груп також мали достовірні відмінності, хоча і були менш виражені (t=2,2; р<0,05) (табл. 2).

При формуванні клітинного моношару, відзначали наявність морфологічних відмінностей між клітинами окістя у щурів першої та другої груп. У всіх спостереженнях у тварин першої групи клітини окістя мали веретеноподібну форму, щільно прилягали одна до одної та розташовувалися паралельно (рис. 5а). У другій групі проліферуючі клітини окістя мали зірчасту (відростчасту) форму з явно вираженими відростками цитоплазми. Періостальні клітини тварин другої групи, на відміну від периостальних клітин тварин першої групи, у процесі культивування утворювали субконфлуент, який мав сітчасту структуру (див. рис. 5б).
При виявленні продукції культивованими клітинами періосту ЛФ позитивну реакцію спостерігали лише у тварин першої групи (рис. 6). Періостальні клітини тварин другої групи ЛФ не продукували.


Дуже показово, що подібна картина спостерігаласт при дослідженні періостальних тканин, отриманих у ході оперативного лікування постраждалих з нарішеннями остеорепарації після переломів кісток. Так, у процесі культивування тканин періосту, взятих у хворих під час оперативного лікування, спостерігали формування моношару, який утворювали гетерогенні клітини. У моношарі спостерігали як довгасті веретеноподібні клітини, так і клітини округлої форми та великих розмірів з цитоплазматичними виростами (рис. 7). За своєю морфологічною картиною моношар, утворений цими клітинами, нагадував моношар, утворений клітинами тварин другої групи. Постановка реакції з BCIP/NBT показала, що серед проліферуючих клітин є поодинокі веретеноподібні клітини, що слабо продукують ЛФ.

При вивченні проліферації клітин ендосту спостерігали картину, схожу на проліферацію клітин періосту. Ендостальні клітини починали проліферувати на 7-8 добу культивування. Кількість проліферуючих ендостальних клітин, як і у випадку з клітинами періосту, на різних рівнях від лінії перелому у двох групах була різною (табл. 3, рис. 8).

У тварин першої групи кількість проліферуючих клітин ендосту протягом усього кісткового фрагмента була приблизно однаковою і коливалася від 35 до 50 у полі зору. У тварин другій групі в області перелому кількість клітин ендосту не перевищувала 12-13 у полі зору, а в зонах віддалених від перелому збільшувалася в 3-4 рази і коливалося від 33 до 49 клітин. У тварин цієї групи достовірні відмінності в кількості клітин, що проліферують, відзначалися тільки між 1-2 рівнями і 3 рівнем (t=2,7, р< 0,02). Відмінностей між 3, 4 і 5 рівнями не було (t=0,4, р> 0,10; t=0,23, р> 0,10 відповідно). Зіставлення кількості клітин на 1 - 5-му рівнях у тварин двох груп показало наявність достовірних відмінностей тільки на перших двох рівнях - t = 4,24, 0,001 для першого рівня і t = 2,74, 0,02 для другого рівня. Морфологія клітин ендосту у тварин першої та другої групи була неоднакова. У тварин першої групи клітин мали округлу форму, характерну для остеобластних клітин (рис 9а). У тварин другої групи клітини, що проліферують, нагадували фібробластоподібні клітини (рис 9б).

При культивуванні ендостальних клітин тварин двох груп було встановлено, що проліферативна активність клітин ендосту у тварин двох груп мала достовірні відмінності. У тварин першої групи проліферативна активність клітин ендосту практично не відрізнялася від контрольних значень (t = 2,7; р <0,05). Проліферативна активність ендостальних клітин тварин другої групи була достовірно вищою за контрольні значення та проліферативну активність клітин ендосту тварин першої групи (t= 5,8; р< 0,001 і t= 5,4; р< 0,001 відповідно) (табл. 4).
До 14-ї доби культивування ендостальні клітини тварин першої групи формували субконфлуентну структуру з остеобластоподібними клітинами, що активно продукують ЛФ (рис. 10а). Ендостальні клітини тварин другої групи до цього часу формували щільний моношар, утворений фібробластоподібними клітинами, що не продукують ЛФ (рис. 10б).

При фазово-контрастній мікроскопії первинних культур мезенхімальних стромальних клітин, виділених з кістково-мозкового каналу травмованих сегментів, було зазначено, що у тварин першої групи кількість МСК, що адгезували до пластику, була приблизно в 10-12 разів більша, ніж у тварин другої групи (рис. 1). Так як здатність адгезувати до пластику мають тільки життєздатні МСК, то дані мікроскопії первинних культур дозволяють говорити про те, що в кістковомозковому каналі під впливом травмуючого агента високої інтенсивності відбувається загибель значної кількості МСК.

За даними МТТ-аналізу проліферативна активність МСК на 14-ту добу культивування у тварин першої групи була в 3,5 рази вище контрольних значень (t= 9,9; р< 0,001), а у тварин 2 групи – у 2,5 раза (t= 7,5). Відмінності у проліферативної активності МСК між групами мали достовірний характер – проліферативна активність МСК тварин першої групи була в 1,4 рази вищою, ніж у тварин другої групи (t=3,2; р<0,01) (табл. 5).

До 14-ї доби культивування МСК КП тварин першої групи формували щільний моношар із клітинами округлої форми, які за своєю морфологією нагадували епітелоїдні клітини (рис. 12а). МСК КП тварин другої групи на той час формували субконфлуент. За своєю структурою субконфлуент, утворений цими клітинами, нагадував субконфлуент, утворений клітинами периоста тварин цієї групи (див. рис. 5б) – одна частина клітин мала веретенообразную форму, а друга частина – зірчасту (рис. 12б).

При зіставленні проліферативної активності різних клітинних джерел остеорепарації у тварин контрольної, першої та другої груп було відзначено, що клітини периоста і МСК, як у контрольній, так і в першій і другій групах, мають більшу проліферативну активність, ніж клітини ендосту. У тварин контрольної групи відмінностей між проліферативною активністю периостальних клітин і МСК не відзначалося (t = 0,63; 0,10), а по відношенню до проліферативної активності ендостальних клітин ці відмінності носили достовірний характер (t = 24,01; р< 0,001 відповідно) (табл. 6, рис. 13).

Ця відмінність зумовлена тим, що клітини періосту та МСК є менш диференційованими, ніж клітини ендоста, які, по своїй суті, є остеобластичними клітинами, що знаходяться на різній стадії диференціювання – від преостеобластів до остеобластів.
Після завдання травми проліферативна активність клітин з різних джерел остеорепарації зростала. Найбільша активація процесів проліферації відзначалася з боку МСК кістково-мозкового каналу. У тварин першої групи проліферативна активність МСК КП була в 2,2 рази вищою, ніж клітин періоста (t = 7,2; р < 0,001) і в 11,3 рази більше ніж клітин ендосту (t = 12,55; р < 0,001). У тварин другої групи проліферативна активність МСК КП була більшою за проліферативну активність клітин періоста в 1,8 раза (t = 5,37; р< 0,001) і в 4,3 рази більше, ніж у клітин ендоста (t = 9,49; р0><. На другому місці проліферативної активності знаходилися клітини періоста. У першій групі їхня активність була в 5,2 рази вищою, ніж активність ендостальних клітин (t = 16,1; р< 0,001), а в другій групі – у 2,4 рази більша (t = 12,7; р< 0,001). У той же час проліферативна активність клітин періосту і МСК КП у тварин першої групи була достовірно вищою, ніж у тварин другої групи (t = 2,19; r = 0,05 і t = 3,19; r = 0,01 відповідно). З боку клітин ендосту, навпаки, більш виражена проліферативна активність відзначалася у тварин другої групи (t = 5,35; r, 0,001).
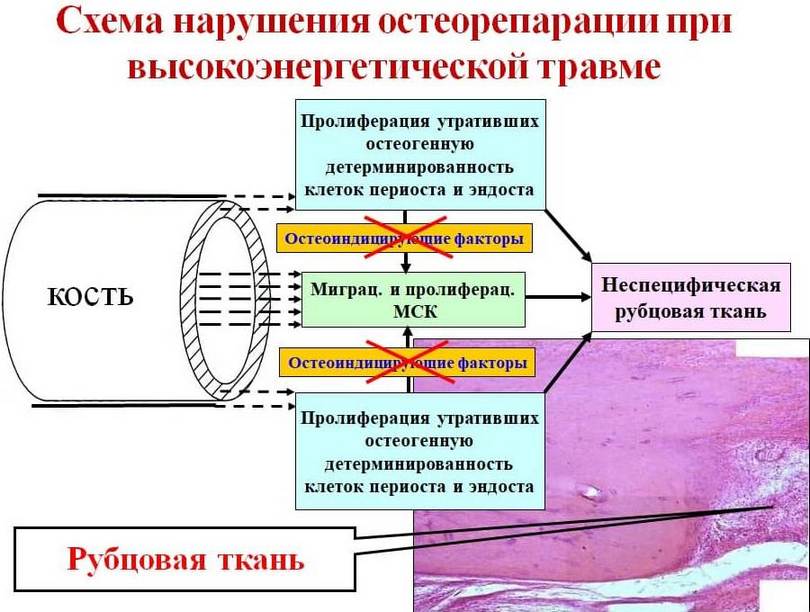
Отримані в результаті досліджень дані свідчать про те, що протягом репаративних процесів у кістковій рані значний вплив має характер травми, а саме інтенсивність, з якою травмуючий агент впливає на кісткову тканину. При травмах низької інтенсивності репаративні процеси, що розвиваються в кістковій рані, призводять до утворення кісткового регенерату та зрощення перелому. При травмах високої інтенсивності у кістковій рані також протікають репаративні процеси, але завершуються формуванням неспецифічної фіброзної тканини (див. рис. 2). Закономірно виникає питання: «Чому після впливу травмуючого агента високої інтенсивності в області перелому не відбувається формування кісткового регенерату?». На думку, відповідь це питання дають результати проведених досліджень.
Отримані при дослідженнях в умовах in vitro дані свідчать про те, що при травмах високої інтенсивності клітини періоста, ендосту та МСК КП зазнають певних морфофункціональних змін. По-перше, при травмі високої інтенсивності в кісткових фрагментах відбувається загибель значної кількості клітинних елементів періоста та ендосту на значній відстані від лінії перелому. Кількість здатних до проліферації клітин періосту у цій галузі скорочується більш ніж у 5 разів, а кількість клітин ендосту зменшується більш ніж у 3,5 рази. Більшість МСК кістковомозкового каналу також втрачає свою життєздатність. На користь цього говорить зменшення більш ніж 10 разів кількості життєздатних МСК. По-друге, проліферативна активність клітин періосту та МСК при травмах високої інтенсивності в 1,2 та 1,4 рази нижча, ніж при травмах низької інтенсивності, а проліферативна активність ендостальних клітин, навпаки, при травмах високої інтенсивності майже у 2 рази вища, ніж при травмах низької інтенсивності. Останнє може бути пояснено дедиференціюванням ендостальних клітин, яке відбувається в результаті впливу на кісткову тканину травмуючого агента високої інтенсивності. На користь дедиференціювання клітин ендосту говорить зміна форми клітин – від округлої, характерної для остеобластних клітин, до веретеноподібної, характерної для фібробластоподібних клітин, які мають більшу проліферативну активність. По-третє, при травмах високої інтенсивності клітини періоста та МСК КП втрачають здатність формувати моношар. Замість моношарів, утворених упорядкованими клітинами, що мають однакову форму, відбувається формування субконфлуентних культур, утворених морфологічно гетерогенними (в т.ч. гіпертрофованими і некротизуючими) клітинами, які мають нехарактерну для клітин періоста і МСК форму (рис. 5, рис. 7). Клітини ендосту, на відміну клітин періоста і МСК, формують щільний моношар, але у освіті моношару беруть участь клітини, мають фібробластоподібну, не характерну для клітин ендосту, форму (див. рис. 10). Крім того, внаслідок впливу травмуючого агента високої інтенсивності, периостальні та ендостальні клітини припиняють продукувати ЛФ.
Також важливо відзначити, що клітинні культури, отримані від групи тварин з низькоенергетичною травмою, морфологічно відмінні від таких, отриманих від тварин з високоенергетичною травмою. Перші мають епітелоїдну морфологію (крім клітин періосту) та є остеобластними лініями (див. рис. 9а, 10а, 12а). Клітини періоста та ендосту тварин першої групи активно продукують ЛФ, що свідчить про їхню остеогенну спрямованість та здатність надавати остеогенну індукцію малодиференційованих клітин кісткового мозку. У той же час, фібробластоїдні клітини тварин другої групи (див. рис. 9б, 10б, 12б) втрачають свою остеогенну детермінованість (про що свідчить відсутність у них продукції ЛФ) і здатність виявляти остеогенну індукцію малодиференційованих клітин кісткового мозку.У роботах останніх років було показано, що в кістковій рані під дією значної кількості біологічно активних речовин відбувається активація та міграція в кісткову рану проліферуючих некоммітованих малодиференційованих МСК КП, які по суті є прото-міофібробластами. При цьому в ділянці перелому локально виділяється значна кількість ТФР-β, накопичується значна кількість специфічного білка фібронектину, і виникають потужні сили натягу екстрацелюлярного матриксу, що відображають механічні властивості матриксу та ремоделюючої активності клітин. Механорецепція між міофібробластами, призводить до значної контракції грануляційної тканини, що утворюється в рані. Після цього поступово відбувається реорганізація грануляційної тканини із запуском процесу апоптозу в міофібробластах та заселенням на їх місце детермінованих та індукованих клітинних елементів.
При переломах на місце міофібробластів, що зазнали апоптозу, приходять МСК з кістковомозкового каналу та остеогенно детерміновані елементи ендосту та періосту. Перистальні та ендостальні клітини, поряд з формуванням кісткового регенерату по периферії, остеоіндукують МСК, що проникають у грануляційну тканину, тим самим створюючи умови для утворення специфічного кісткового регенерату у всій товщі кісткової рани (рис. 12). У тих випадках, коли, з будь-яких причин (у нашому випадку внаслідок впливу травми високої інтенсивності), клітини періосту та ендосту втрачають свою остеогенну детермінацію, остеоіндукуючого впливу з їхнього боку на МСК, що знаходяться в грануляційній тканині, не відбувається. В результаті цього при реорганізації грануляційної тканини кісткової рани на місце міофібробластів, що зазнали апоптозу, з кістковомозкового каналу, гаверсових каналів і області периоста мігрують морфологічно змінені і втратили властиві їм остеогенні властивості фібробластоподібні клітини, які сприяють утворенню (рис 13).


Таким чином, узагальнюючи все вище сказане, можна говорити про те, що однією з основних причин, що порушують перебіг репаративного остеогенезу, при травмах високої інтенсивності є морфофункціональні зміни клітинних джерел остеорепарації. Останні характеризуються дедиференціюванням остеогенно детермінованих клітин періоста та ендосту, що призводить до розвитку остеогенної недостатності, яка була описана В.Г. Гололобовим із співавт. на підставі емпіричних висновків та підтверджена нашими експериментальними дослідженнями. Механізми, що призводять до морфофункціональних змін клітинних джерел остеорепарації, нам не відомі та потребують окремого вивчення.
Проблемі лікування посттравматичної кісткової дисрегенерації присвячено значну кількість робіт, у яких розглядаються як хірургічні, так і нехірургічні методи стимуляції репаративного остеогенезу. Однак, незважаючи на досить високий відсоток успішного лікування хворих із порушенням процесів консолідації кісткових уламків, у 4,8 – 15,4% хворих досягти позитивних результатів не вдається. Найбільші труднощі при лікуванні порушень остеорепарації виникають після травм, отриманих внаслідок впливу високоенергетичного агента, що травмує. За даними різних джерел, у 10,8 – 13,7% хворих з високоенергетичними травмами нижніх кінцівок переломи не зросталися протягом 1,5 – 3 років. Відомо, що формування кісткової мозолі в області перелому відбувається завдяки клітинам-попередникам періосту, ендосту та мезенхімальним стромальним (стволовим) клітинам кістковомозкової порожнини (МСК КП). Одним з визначальних механізмів формування кісткової мозолі є остеогенне диференціювання клітин-попередників, що проліферують у кісткову рану. Дослідження останніх років показали, що диференціація МСК за остеобластичним типом є складним, багатогранним процесом і здійснюється під контролем факторів транскрипції, клітинних і матриксних взаємодій, системних і місцевих факторів. На процеси остеогенної диференціації значний вплив дуже багато місцевих факторів, переважна більшість яких продукується клітинами остеобластической лінії. Але, незважаючи на те, що в процесі остеогенного диференціювання клітин-попередників, що мігрують у кісткову рану, провідне значення відіграють паракринні фактори, що продукуються вже мають остеогенну спрямованість клітинами периоста і ендоста, робіт, присвячених вивченню змін морфофункціонального стану клітинних джерел остеорепарації в залежності від механізму травм знайти не вдалося.
У зв'язку з цим було проведено вивчення ролі змін морфофункціонального стану клітинних джерел остеорепарації (періосту, ендосту та мезенхімальних стромальних клітин кістково-мозкової порожнини (МСК КП) у порушенні репаративного остеогенезу. Для цього були проведені експериментальні та клінічні дослідження. Вивчали морфологічну картину. остеорепарації (періоста, ендосту та МСК КП).
Експериментальні дослідження проводилися in vivo та in vitro на 80 статевозрілих білих безпородних щурах самцях. Тварин підрозділили на три групи – контрольну, першу та другу. Контрольну групу утворили десять тварин. У цих тварин за допомогою МТТ-аналізу вивчали проліферативну активність клітин періоста, ендосту та МСК КП. Для чого після виведення з експерименту у інтактних тварин в асептичних умовах здійснювали відчленування обох гомілок і виділення з м'яких тканин гомілкових кісток. Берцеві кістки поміщали у спеціальний консервант та витримували в ньому протягом доби. Потім у стерильних умовах ножицями відсікали метаепіфізарні області, залишаючи для подальшого дослідження лише діафізарну частину кістки. Для визначення проліферативної активності МСК КП у стерильних умовах з кістковомозкового каналу за допомогою інсулінового шприца та фізіологічного розчину вимивали кістковомозковий вміст у чашки Петрі з культуральним середовищем. Для спрощення цієї процедури діафізарну частину кістки розтинали поперечно навпіл. Половина кісткових фрагментів розсікалася поздовжньо очними ножицями, ретельно відмивали від залишків кістковомозкового вмісту фізіологічним розчином і поміщали в чашки Петрі з колагеновою підкладкою і живильним середовищем таким чином, щоб з колагеновою підкладкою контактувала тільки окістя (мал. Таке становище кісткових фрагментів у чашках Петрі створювало умови для проліферації лише клітин окістя. Половину кісткових фрагментів, що залишилася, за допомогою скальпеля, ретельно очищали від окістя, промивали фізіологічним розчином і поміщали в чашки Петрі з колагеновою підкладкою і живильним середовищем таким чином, щоб з колагеновою підкладкою контактувала кісткова тканина (рис. 1 б). Це створювало умови для проліферації лише клітин власне кісткової тканини (клітин ендосту).
У тварин першої (35 щурів) та другої (35 щурів) груп моделювали низько- та високоенергетичні переломи гомілкової кістки. При моделюванні низькоенергетичних травм (перша група) в асептичних умовах під ефірним наркозом в області середньої третини гомілки скальпелем здійснювали розріз шкіри і виділяли гомілкову кістку. Вузьким долотом виконували остеотомію кістки в середній третині, після чого зашивали рану наглухо. При моделюванні високоенергетичних переломів (2 група) під ефірним наркозом області середньої третини гомілки за допомогою пружинного пристрою завдавали дозований удар, сила якого становила в середньому 2,55±0,04 кг×м/с. У 10 тварин (по 5 тварин кожної групи) травмовані сегменти фіксували параосально з використанням голок для внутрішньом'язових ін'єкцій та поліхлорвінілових трубок для внутрішньовенних інфузій. Виведення цих тварин з експерименту здійснювали на 7 добу після травми. Після виведення з експерименту у тварин в асептичних умовах відчленовували травмований сегмент, звільняли його від шкіри, фіксували в 10% розчині нейтрального формаліну, декальцинували і фарбували гематоксиліном та еозином. Отримані гістологічні препарати вивчали у світловому мікроскопі. Шістдесят тварин (по 30 тварин кожної групи) було виведено з експерименту через 1 годину після травми. Після виведення з експерименту у тварин в асептичних умовах відчленовували травмований сегмент, кісткові фрагменти акуратно виділяли з м'яких тканин та поміщали у флакон зі стерильним консервантом. Кісткові фрагменти витримували у флаконі з консервантом протягом однієї доби. Потім з кістковими фрагментами кожної з тварин виконували ті ж маніпуляції, що і у тварин контрольної групи і культивували в 22-інкубаторі за стандартних умов. На 5 – 8-му добу культивування за допомогою фазово-контрастної мікроскопії здійснювали підрахунок проліферуючих від кісткових фрагментів клітин періоста та ендосту. Клітини підраховували у зору мікроскопа при 100-кратном збільшенні на п'яти рівнях від лінії перелому. Як критерій, що визначає рівень підрахунку, використовували поле зору мікроскопа. На 14-ту добу культивування в 45 чашках Петрі кожної групи (по 15 чашок для кожної з клітинних культур) методом МТТ-аналізу визначали проліферативну активність клітин періоста, ендосту та МСК КП. В решті 45-ти чашок Петрі кожної групи (по 15 чашок для кожної з клітинних культур), після формування моношару за допомогою фазово-контрастної мікроскопії оцінювали морфологічний стан проліферуючих клітин окістя, ендосту та МСК КП. У моношарі культур періоста та ендосту за допомогою реакції з BCIP/NBT Liquid Substrate System (Sigma, США) визначали наявність або відсутність продукції клітинами лужної фосфатази (ЩФ), що дозволяло судити про наявність або відсутність остеогенної детермінованості (спрямованості) клітин.
Клінічні дослідження були проведені у 5-ти хворих із тривалим незрощенням переломів довгих кісток нижніх кінцівок. У 2-х хворих були незрощені переломи стегнової кістки, у 3-х хворих - великогомілкової кістки. Тривалість порушення цілісності кісткової тканини у хворих була від 11 місяців до 2 років. Під час оперативного лікування у хворих із проксимального кісткового фрагмента в області перелому здійснювали забір періостальних тканин розміром 0,5'0,5 см. Біопсійний матеріал поміщали у флакон із консервантом та витримували протягом доби. Потім у стерильних умовах ділянки периоста витягали з консерванту і поміщали в чашки Петрі з живильним середовищем і культивували в <2>2-інкубаторі за стандартних умов. Після появи клітинної проліферації періостальні тканини витягувалися, а клітини, що прикріпилися до пластику, культивувалися до формування моношару. За допомогою фазово-контрастної мікроскопії оцінювали морфологічний стан проліферуючих клітин періосту, а за допомогою реакції на ЛФ – їхню остеогенну детермінованість.
Фотодокументування клітинних культур та гістопрепаратів здійснювали за допомогою інвертованого мікроскопа Leica DMIL, робочої станції з обробки зображень Leica QWin500 Standart (версія 2.3, сер. № 3069) та відеокамери Sanyo TK-C1380 (Німеччина). Отриманий у результаті досліджень цифровий матеріал піддавали статистичній обробці.
При вивченні гістоморфологічних препаратів області кісткової рани на 7-му добу після травми було зазначено, що стан кісткових фрагментів та зрощення переломів у тварин першої та другої груп було по-різному. У тварин першої групи кісткові фрагменти мали значну кількість клітинних елементів із добре фарбованими ядрами. В області кісткової рани відбувалося утворення примітивних кісткових балок, характерних для формування кісткової мозолі (рис. 2а). У другій групі кількість клітинних елементів у кістковій тканині уламків було набагато менше, ніж у першій групі. Вони розташовувалися переважно у віддалених від лінії перелому ділянках кісткової тканини. У кістки, що прилягає безпосередньо до лінії перелому, зустрічалися поодинокі клітини із слабо профарбованими ядрами. Ці зміни морфологічно нагадували картину остеонекрозу. В області перелому репаративні процеси виявлялися у вигляді формування грубоволокнистої ретикулярної сполучної тканини, що заповнює кісткову рану (див. рис. 2б).
Таким чином, дані гістоморфологічних досліджень свідчать про те, що інтенсивність впливу на кістку травмуючого агента впливає стан кісткової тканини і протягом зрощення перелому. При низькоенергетичних травмах у кістковій рані репаративні процеси протікають із формуванням специфічного кісткового регенерату, а при високоенергетичних травмах репаративні процеси обмежуються утворенням неспецифічного рубцевого регенерату.
При культивуванні клітин окістя активна клітинна проліферація починалася на 5-6 добу. У тварин першої групи клітини окістя проліферували рівномірно вздовж всього кісткового фрагмента (рис 3а). У тварин другої групи проліферація клітин окістя вздовж кісткового фрагмента була нерівномірною. У зоні, що прилягає до лінії перелому, проліферували поодинокі клітини. Активна клітинна проліферація відзначалася у зоні, найбільш віддаленій від лінії перелому (рис. 3б).
Результати підрахунку кількості клітин періосту у тварин 1 та 2 груп представлені у таблиці 1 та на діаграмі (рис. 4).
Наведені дані свідчать про те, що у тварин першої групи кількість проліферуючих клітин була однаковою по всьому фрагменту кістки і становила 27-29 клітин у полі зору. У тварин другої групи кількість проліферуючих клітин у зоні перелому не перевищувала 6 у полі зору. У більш віддалених від лінії зламу зонах кількість проліферуючих клітин поступово зростала від 17 до 54 клітин. Відмінності в кількості клітин між рівнями від лінії перелому у тварин другої групи були вірогідними (між рівнями 1, 2 і 3 - t=2,39, p<0,05; між рівнями 3 і 4 - t=2,3, p<0,05; між рівнями 4 і 5 - t=2,2, p<0,05).
Зіставлення кількості клітин на 1 – 5-му рівнях у тварин двох груп показало, що у тварин першої групи, на перших трьох рівнях кількість клітин, що проліферують, значно перевищувала таку у тварин другої групи (р< 0,002 і р < 0,05). На четвертому рівні кількість проліферуючих клітин окістя в обох групах була приблизно однаковою (р> 0,10), а на п'ятому рівні від лінії перелому відмінності мали зворотну залежність. У цій галузі кількість проліферуючих клітин у другій групі була достовірно більшою, ніж у першій групі (р<0,05).
У процесі культивування проліферативна активність клітин періосту у тварин двох груп була вищою за контрольне значення. Проліферативна активність клітин періоста в першій групі була в 1,6 рази (t= 7,8; р< 0,001), а в другій в 1,4 раза (t= 7,7; р< 0,001) більше контрольних значень. Відмінності між проліферативною активністю клітин періоста першої та другої груп також мали достовірні відмінності, хоча і були менш виражені (t=2,2; р<0,05) (табл. 2).
При формуванні клітинного моношару, відзначали наявність морфологічних відмінностей між клітинами окістя у щурів першої та другої груп. У всіх спостереженнях у тварин першої групи клітини окістя мали веретеноподібну форму, щільно прилягали одна до одної та розташовувалися паралельно (рис. 5а). У другій групі проліферуючі клітини окістя мали зірчасту (відростчасту) форму з явно вираженими відростками цитоплазми. Періостальні клітини тварин другої групи, на відміну від периостальних клітин тварин першої групи, у процесі культивування утворювали субконфлуент, який мав сітчасту структуру (див. рис. 5б).
При виявленні продукції культивованими клітинами періосту ЛФ позитивну реакцію спостерігали лише у тварин першої групи (рис. 6). Періостальні клітини тварин другої групи ЛФ не продукували.
Дуже показово, що подібна картина спостерігаласт при дослідженні періостальних тканин, отриманих у ході оперативного лікування постраждалих з нарішеннями остеорепарації після переломів кісток. Так, у процесі культивування тканин періосту, взятих у хворих під час оперативного лікування, спостерігали формування моношару, який утворювали гетерогенні клітини. У моношарі спостерігали як довгасті веретеноподібні клітини, так і клітини округлої форми та великих розмірів з цитоплазматичними виростами (рис. 7). За своєю морфологічною картиною моношар, утворений цими клітинами, нагадував моношар, утворений клітинами тварин другої групи. Постановка реакції з BCIP/NBT показала, що серед проліферуючих клітин є поодинокі веретеноподібні клітини, що слабо продукують ЛФ.
При вивченні проліферації клітин ендосту спостерігали картину, схожу на проліферацію клітин періосту. Ендостальні клітини починали проліферувати на 7-8 добу культивування. Кількість проліферуючих ендостальних клітин, як і у випадку з клітинами періосту, на різних рівнях від лінії перелому у двох групах була різною (табл. 3, рис. 8).
У тварин першої групи кількість проліферуючих клітин ендосту протягом усього кісткового фрагмента була приблизно однаковою і коливалася від 35 до 50 у полі зору. У тварин другій групі в області перелому кількість клітин ендосту не перевищувала 12-13 у полі зору, а в зонах віддалених від перелому збільшувалася в 3-4 рази і коливалося від 33 до 49 клітин. У тварин цієї групи достовірні відмінності в кількості клітин, що проліферують, відзначалися тільки між 1-2 рівнями і 3 рівнем (t=2,7, р< 0,02). Відмінностей між 3, 4 і 5 рівнями не було (t=0,4, р> 0,10; t=0,23, р> 0,10 відповідно). Зіставлення кількості клітин на 1 - 5-му рівнях у тварин двох груп показало наявність достовірних відмінностей тільки на перших двох рівнях - t = 4,24, 0,001 для першого рівня і t = 2,74, 0,02 для другого рівня. Морфологія клітин ендосту у тварин першої та другої групи була неоднакова. У тварин першої групи клітин мали округлу форму, характерну для остеобластних клітин (рис 9а). У тварин другої групи клітини, що проліферують, нагадували фібробластоподібні клітини (рис 9б).
При культивуванні ендостальних клітин тварин двох груп було встановлено, що проліферативна активність клітин ендосту у тварин двох груп мала достовірні відмінності. У тварин першої групи проліферативна активність клітин ендосту практично не відрізнялася від контрольних значень (t = 2,7; р <0,05). Проліферативна активність ендостальних клітин тварин другої групи була достовірно вищою за контрольні значення та проліферативну активність клітин ендосту тварин першої групи (t= 5,8; р< 0,001 і t= 5,4; р< 0,001 відповідно) (табл. 4).
До 14-ї доби культивування ендостальні клітини тварин першої групи формували субконфлуентну структуру з остеобластоподібними клітинами, що активно продукують ЛФ (рис. 10а). Ендостальні клітини тварин другої групи до цього часу формували щільний моношар, утворений фібробластоподібними клітинами, що не продукують ЛФ (рис. 10б).
При фазово-контрастній мікроскопії первинних культур мезенхімальних стромальних клітин, виділених з кістково-мозкового каналу травмованих сегментів, було зазначено, що у тварин першої групи кількість МСК, що адгезували до пластику, була приблизно в 10-12 разів більша, ніж у тварин другої групи (рис. 1). Так як здатність адгезувати до пластику мають тільки життєздатні МСК, то дані мікроскопії первинних культур дозволяють говорити про те, що в кістковомозковому каналі під впливом травмуючого агента високої інтенсивності відбувається загибель значної кількості МСК.
За даними МТТ-аналізу проліферативна активність МСК на 14-ту добу культивування у тварин першої групи була в 3,5 рази вище контрольних значень (t= 9,9; р< 0,001), а у тварин 2 групи – у 2,5 раза (t= 7,5). Відмінності у проліферативної активності МСК між групами мали достовірний характер – проліферативна активність МСК тварин першої групи була в 1,4 рази вищою, ніж у тварин другої групи (t=3,2; р<0,01) (табл. 5).
До 14-ї доби культивування МСК КП тварин першої групи формували щільний моношар із клітинами округлої форми, які за своєю морфологією нагадували епітелоїдні клітини (рис. 12а). МСК КП тварин другої групи на той час формували субконфлуент. За своєю структурою субконфлуент, утворений цими клітинами, нагадував субконфлуент, утворений клітинами периоста тварин цієї групи (див. рис. 5б) – одна частина клітин мала веретенообразную форму, а друга частина – зірчасту (рис. 12б).
При зіставленні проліферативної активності різних клітинних джерел остеорепарації у тварин контрольної, першої та другої груп було відзначено, що клітини периоста і МСК, як у контрольній, так і в першій і другій групах, мають більшу проліферативну активність, ніж клітини ендосту. У тварин контрольної групи відмінностей між проліферативною активністю периостальних клітин і МСК не відзначалося (t = 0,63; 0,10), а по відношенню до проліферативної активності ендостальних клітин ці відмінності носили достовірний характер (t = 24,01; р< 0,001 відповідно) (табл. 6, рис. 13).
Ця відмінність зумовлена тим, що клітини періосту та МСК є менш диференційованими, ніж клітини ендоста, які, по своїй суті, є остеобластичними клітинами, що знаходяться на різній стадії диференціювання – від преостеобластів до остеобластів.
Після завдання травми проліферативна активність клітин з різних джерел остеорепарації зростала. Найбільша активація процесів проліферації відзначалася з боку МСК кістково-мозкового каналу. У тварин першої групи проліферативна активність МСК КП була в 2,2 рази вищою, ніж клітин періоста (t = 7,2; р < 0,001) і в 11,3 рази більше ніж клітин ендосту (t = 12,55; р < 0,001). У тварин другої групи проліферативна активність МСК КП була більшою за проліферативну активність клітин періоста в 1,8 раза (t = 5,37; р< 0,001) і в 4,3 рази більше, ніж у клітин ендоста (t = 9,49; р0><. На другому місці проліферативної активності знаходилися клітини періоста. У першій групі їхня активність була в 5,2 рази вищою, ніж активність ендостальних клітин (t = 16,1; р< 0,001), а в другій групі – у 2,4 рази більша (t = 12,7; р< 0,001). У той же час проліферативна активність клітин періосту і МСК КП у тварин першої групи була достовірно вищою, ніж у тварин другої групи (t = 2,19; r = 0,05 і t = 3,19; r = 0,01 відповідно). З боку клітин ендосту, навпаки, більш виражена проліферативна активність відзначалася у тварин другої групи (t = 5,35; r, 0,001).
Отримані в результаті досліджень дані свідчать про те, що протягом репаративних процесів у кістковій рані значний вплив має характер травми, а саме інтенсивність, з якою травмуючий агент впливає на кісткову тканину. При травмах низької інтенсивності репаративні процеси, що розвиваються в кістковій рані, призводять до утворення кісткового регенерату та зрощення перелому. При травмах високої інтенсивності у кістковій рані також протікають репаративні процеси, але завершуються формуванням неспецифічної фіброзної тканини (див. рис. 2). Закономірно виникає питання: «Чому після впливу травмуючого агента високої інтенсивності в області перелому не відбувається формування кісткового регенерату?». На думку, відповідь це питання дають результати проведених досліджень.
Отримані при дослідженнях в умовах in vitro дані свідчать про те, що при травмах високої інтенсивності клітини періоста, ендосту та МСК КП зазнають певних морфофункціональних змін. По-перше, при травмі високої інтенсивності в кісткових фрагментах відбувається загибель значної кількості клітинних елементів періоста та ендосту на значній відстані від лінії перелому. Кількість здатних до проліферації клітин періосту у цій галузі скорочується більш ніж у 5 разів, а кількість клітин ендосту зменшується більш ніж у 3,5 рази. Більшість МСК кістковомозкового каналу також втрачає свою життєздатність. На користь цього говорить зменшення більш ніж 10 разів кількості життєздатних МСК. По-друге, проліферативна активність клітин періосту та МСК при травмах високої інтенсивності в 1,2 та 1,4 рази нижча, ніж при травмах низької інтенсивності, а проліферативна активність ендостальних клітин, навпаки, при травмах високої інтенсивності майже у 2 рази вища, ніж при травмах низької інтенсивності. Останнє може бути пояснено дедиференціюванням ендостальних клітин, яке відбувається в результаті впливу на кісткову тканину травмуючого агента високої інтенсивності. На користь дедиференціювання клітин ендосту говорить зміна форми клітин – від округлої, характерної для остеобластних клітин, до веретеноподібної, характерної для фібробластоподібних клітин, які мають більшу проліферативну активність. По-третє, при травмах високої інтенсивності клітини періоста та МСК КП втрачають здатність формувати моношар. Замість моношарів, утворених упорядкованими клітинами, що мають однакову форму, відбувається формування субконфлуентних культур, утворених морфологічно гетерогенними (в т.ч. гіпертрофованими і некротизуючими) клітинами, які мають нехарактерну для клітин періоста і МСК форму (рис. 5, рис. 7). Клітини ендосту, на відміну клітин періоста і МСК, формують щільний моношар, але у освіті моношару беруть участь клітини, мають фібробластоподібну, не характерну для клітин ендосту, форму (див. рис. 10). Крім того, внаслідок впливу травмуючого агента високої інтенсивності, периостальні та ендостальні клітини припиняють продукувати ЛФ.
Також важливо відзначити, що клітинні культури, отримані від групи тварин з низькоенергетичною травмою, морфологічно відмінні від таких, отриманих від тварин з високоенергетичною травмою. Перші мають епітелоїдну морфологію (крім клітин періосту) та є остеобластними лініями (див. рис. 9а, 10а, 12а). Клітини періоста та ендосту тварин першої групи активно продукують ЛФ, що свідчить про їхню остеогенну спрямованість та здатність надавати остеогенну індукцію малодиференційованих клітин кісткового мозку. У той же час, фібробластоїдні клітини тварин другої групи (див. рис. 9б, 10б, 12б) втрачають свою остеогенну детермінованість (про що свідчить відсутність у них продукції ЛФ) і здатність виявляти остеогенну індукцію малодиференційованих клітин кісткового мозку.У роботах останніх років було показано, що в кістковій рані під дією значної кількості біологічно активних речовин відбувається активація та міграція в кісткову рану проліферуючих некоммітованих малодиференційованих МСК КП, які по суті є прото-міофібробластами. При цьому в ділянці перелому локально виділяється значна кількість ТФР-β, накопичується значна кількість специфічного білка фібронектину, і виникають потужні сили натягу екстрацелюлярного матриксу, що відображають механічні властивості матриксу та ремоделюючої активності клітин. Механорецепція між міофібробластами, призводить до значної контракції грануляційної тканини, що утворюється в рані. Після цього поступово відбувається реорганізація грануляційної тканини із запуском процесу апоптозу в міофібробластах та заселенням на їх місце детермінованих та індукованих клітинних елементів.
При переломах на місце міофібробластів, що зазнали апоптозу, приходять МСК з кістковомозкового каналу та остеогенно детерміновані елементи ендосту та періосту. Перистальні та ендостальні клітини, поряд з формуванням кісткового регенерату по периферії, остеоіндукують МСК, що проникають у грануляційну тканину, тим самим створюючи умови для утворення специфічного кісткового регенерату у всій товщі кісткової рани (рис. 12). У тих випадках, коли, з будь-яких причин (у нашому випадку внаслідок впливу травми високої інтенсивності), клітини періосту та ендосту втрачають свою остеогенну детермінацію, остеоіндукуючого впливу з їхнього боку на МСК, що знаходяться в грануляційній тканині, не відбувається. В результаті цього при реорганізації грануляційної тканини кісткової рани на місце міофібробластів, що зазнали апоптозу, з кістковомозкового каналу, гаверсових каналів і області периоста мігрують морфологічно змінені і втратили властиві їм остеогенні властивості фібробластоподібні клітини, які сприяють утворенню (рис 13).
Таким чином, узагальнюючи все вище сказане, можна говорити про те, що однією з основних причин, що порушують перебіг репаративного остеогенезу, при травмах високої інтенсивності є морфофункціональні зміни клітинних джерел остеорепарації. Останні характеризуються дедиференціюванням остеогенно детермінованих клітин періоста та ендосту, що призводить до розвитку остеогенної недостатності, яка була описана В.Г. Гололобовим із співавт. на підставі емпіричних висновків та підтверджена нашими експериментальними дослідженнями. Механізми, що призводять до морфофункціональних змін клітинних джерел остеорепарації, нам не відомі та потребують окремого вивчення.