The problem of treating post-traumatic bone dysregeneration is the subject of a significant number of works, which consider both surgical and non-surgical methods of stimulating reparative osteogenesis. However, despite a fairly high percentage of successful treatment of patients with impaired consolidation of bone fragments, in 4.8 - 15.4% of patients it is not possible to achieve positive results. The greatest difficulties in the treatment of osteoreparation disorders arise after injuries resulting from exposure to a high-energy traumatic agent. According to various sources, in 10.8 - 13.7% of patients with high-energy injuries of the lower extremities, fractures did not heal within 1.5 - 3 years. It is known that the formation of callus in the fracture area occurs due to the progenitor cells of the periosteum, endosteum and mesenchymal stromal (stem) cells of the bone marrow cavity (MSCs). One of the determining mechanisms of callus formation is osteogenic differentiation of precursor cells proliferating into the bone wound. Recent studies have shown that differentiation of MSCs according to the osteoblastic type is a complex, multifaceted process and is carried out under the control of transcription factors, cellular and matrix interactions, systemic and local factors. The processes of osteogenic differentiation are significantly influenced by a large number of local factors, the vast majority of which are produced by cells of the osteoblastic line. But, despite the fact that in the process of osteogenic differentiation of precursor cells migrating into a bone wound, paracrine factors produced by periosteal and endosteal cells that already have an osteogenic orientation play a leading role, studies devoted to the study of changes in the morphofunctional state of cellular sources of osteoreparation depending on the mechanism of injury and the role of these changes in the development of dysregeneration could not be found in the available literature.
In this regard, a study was carried out of the role of changes in the morphofunctional state of cellular sources of osteoreparation (periosteum, endosteum and mesenchymal stromal cells of the bone marrow cavity (MCCs) in the disruption of reparative osteogenesis. For this purpose, experimental and clinical studies were carried out. The morphological picture of the bone wound area and the morphofunctional state of cellular sources of osteoreparation (periosteum, endosteum and MSCs) were studied KP).
Experimental studies were carried out in vivo and in vitro on 80 mature white outbred male rats. The animals were divided into three groups – control, first and second. Ten animals formed the control group. In these animals, the proliferative activity of periosteal, endosteal and PC MSCs was studied using MTT analysis. For this purpose, after removal from the experiment in intact animals, both legs were separated under aseptic conditions and the tibia bones were isolated from the soft tissues. The tibia bones were placed in a special preservative and kept in it for 24 hours. Then, under sterile conditions, the metaepiphyseal regions were cut off with scissors, leaving only the diaphyseal part of the bone for further examination. To determine the proliferative activity of PC MSCs under sterile conditions, the bone marrow contents were washed out of the bone marrow canal using an insulin syringe and physiological solution into Petri dishes with a culture medium. To simplify this procedure, the diaphyseal part of the bone was cut transversely in half. Half of the bone fragments were dissected longitudinally with ophthalmic scissors, thoroughly washed from the remaining bone marrow contents with physiological solution and placed in Petri dishes with a collagen substrate and nutrient medium so that only the periosteum was in contact with the collagen substrate (Fig. 1a). This position of bone fragments in Petri dishes created conditions for the proliferation of only periosteal cells. The remaining half of the bone fragments were carefully cleared from the periosteum using a scalpel, washed with saline and placed in Petri dishes with a collagen substrate and nutrient medium so that the bone tissue was in contact with the collagen substrate (Fig. 1 b). This created conditions for the proliferation of only cells of the bone tissue itself (endosteal cells).

Petri dishes with bone marrow contents and bone fragments were placed in a CO2-incubator and cultured under standard conditions (temperature 37°C and 5% CO2 content) with the nutrient medium changed every third day. After the start of cell proliferation in Petri dishes with bone fragments, the latter were carefully removed, and the cells attached to the collagen substrate continued to be cultured. MTT analysis of the proliferative activity of periosteal, endosteal and PC MSCs was carried out on the 14th day of cultivation. The results obtained were accepted as control ones.
Low- and high-energy fractures of the tibia were modeled in animals of the first (35 rats) and second (35 rats) groups. When simulating low-energy injuries (first group) under aseptic conditions, under ether anesthesia in the area of the middle third of the leg, a skin incision was made with a scalpel and the tibia was isolated. An osteotomy of the bone in the middle third was performed with a narrow chisel, after which the wound was sutured tightly. When simulating high-energy fractures (group 2), under ether anesthesia, a dosed blow was applied in the area of the middle third of the leg using a spring device, the force of which averaged 2.55±0.04 kg×m/s. In 10 animals (5 animals in each group), the injured segments were fixed paraosseously using needles for intramuscular injections and polyvinyl chloride tubes for intravenous infusions. These animals were removed from the experiment on the 7th day after injury. After the animals were removed from the experiment, the injured segment was separated under aseptic conditions, freed from the skin, fixed in a 10% solution of neutral formaldehyde, decalcified and stained with hematoxylin and eosin. The resulting histological preparations were studied under a light microscope. Sixty animals (30 animals from each group) were removed from the experiment 1 hour after injury. After the animals were removed from the experiment, the injured segment was separated under aseptic conditions, bone fragments were carefully isolated from the soft tissues and placed in a bottle with a sterile preservative. The bone fragments were kept in a bottle with a preservative for one day. Then, the same manipulations were performed with the bone fragments of each of the animals as in the animals of the control group and were cultured in a CO2 incubator under standard conditions. On days 5–8 of cultivation, periosteal and endosteal cells proliferating from bone fragments were counted using phase-contrast microscopy. Cells were counted in the field of view of the microscope at 100× magnification at five levels from the fracture line. The field of view of the microscope was used as a criterion to determine the level of counting. On the 14th day of cultivation in 45 Petri dishes of each group (15 dishes for each cell culture), the proliferative activity of periosteal, endosteal and PC MSCs was determined using MTT analysis. In the remaining 45 Petri dishes of each group (15 dishes for each cell culture), after the formation of a monolayer, the morphological state of proliferating cells of the periosteum, endosteum and PC MSCs was assessed using phase-contrast microscopy. In a monolayer of periosteal and endosteal cultures, the presence or absence of alkaline phosphatase (ALP) production by cells was determined using a reaction with BCIP/NBT Liquid Substrate System (Sigma, USA), which made it possible to judge the presence or absence of osteogenic determination (directionality) of cells.
Clinical studies were conducted in 5 patients with long-term nonunion of fractures of the long bones of the lower extremities. 2 patients had ununited fractures of the femur, and 3 patients had ununited fractures of the tibia. The duration of damage to the integrity of bone tissue in these patients ranged from 11 months to 2 years. During surgical treatment in patients, periosteal tissue measuring 0.5 x 0.5 cm was taken from the proximal bone fragment in the area of the fracture. The biopsy material was placed in a bottle with a preservative and kept for 24 hours. Then, under sterile conditions, sections of the periosteum were removed from the preservative and placed in Petri dishes with a nutrient medium and cultured in a CO2 incubator under standard conditions. After the appearance of cell proliferation, the periosteal tissues were removed, and the cells attached to the plastic were cultured until a monolayer was formed. Using phase-contrast microscopy, the morphological state of proliferating periosteal cells was assessed, and their osteogenic determination was assessed using the reaction to alkaline phosphatase.
Photo documentation of cell cultures and histo-preparations was carried out using a Leica DMIL inverted microscope, a Leica QWin500 Standart image processing workstation (version 2.3, sir. No. 3069) and a Sanyo TK-C1380 video camera (Germany). The digital material obtained as a result of the research was subjected to statistical processing.
When studying histomorphological preparations of the bone wound area on the 7th day after the injury, it was noted that the condition of the bone fragments and the healing of fractures in animals of the first and second groups was different. In animals of the first group, bone fragments had a significant number of cellular elements with well-stained nuclei. In the area of the bone wound, primitive bone beams formed, characteristic of the processes of callus formation (Fig. 2a). In the second group, the number of cellular elements in the bone tissue of the fragments was many times less than in the first group. They were located predominantly in areas of bone tissue remote from the fracture line. In the bone immediately adjacent to the fracture line, there were single cells with weakly stained nuclei. These changes morphologically resembled the picture of osteonecrosis. In the area of the fracture, reparative processes manifested themselves in the form of the formation of coarse-fibrous reticular connective tissue filling the bone wound (see Fig. 2b).

Thus, data from histomorphological studies indicate that the intensity of the traumatic agent affecting the bone affects the condition of the bone tissue and the course of fracture healing. With low-energy injuries in a bone wound, reparative processes occur with the formation of a specific bone regenerate, and with high-energy injuries, reparative processes are limited to the formation of nonspecific scar regenerate.
When culturing periosteum cells active cell proliferation began on the 5-6th day. In animals of the first group, periosteal cells proliferated evenly along the entire bone fragment (Fig. 3a). In animals of the second group, the proliferation of periosteal cells along the bone fragment was uneven. In the area adjacent to the fracture line, single cells proliferated. Active cell proliferation was observed in the area furthest from the fracture line (Fig. 3b).

The results of counting the number of periosteal cells in animals of groups 1 and 2 are presented in Table 1 and in the diagram (Fig. 4).

The presented data indicate that in animals of the first group the number of proliferating cells was the same throughout the entire bone fragment and amounted to 27-29 cells in the field of view. In the second group of animals, the number of proliferating cells in the fracture area did not exceed 6 in the field of view. In zones more distant from the fracture line, the number of proliferating cells gradually increased from 17 to 54 cells. The differences in the number of cells between levels from the fracture line in animals of the second group were significant (between levels 1, 2 and 3 - t=2.39, p<0.05; between levels 3 and 4 - t=2.3, p<0.05; between levels 4 and 5 - t=2.2, p<0.05).
A comparison of the number of cells at levels 1–5 in animals of the two groups showed that in animals of the first group, at the first three levels, the number of proliferating cells significantly exceeded that in animals of the second group (p< 0.002 andp < 0.05). At the fourth level, the number of proliferating periosteal cells in both groups was approximately the same (p> 0.10), and at the fifth level, the differences had an inverse relationship with the fracture line. In this area, the number of proliferating cells in the second group was significantly greater than in the first group (p<0.05).
During the cultivation process, the proliferative activity of periosteal cells in animals of the two groups was higher than the control value. The proliferative activity of periosteal cells in the first group was 1.6 times (t= 7.8; p< 0.001), and in the second group it was 1.4 times (t= 7.7; p< 0.001) higher than control values. The differences between the proliferative activity of periosteal cells of the first and second groups also had significant differences, although they were less pronounced (t = 2.2; p< 0.05) (Table 2).

During the formation of a cell monolayer, the presence of morphological differences between the cells of the periosteum in rats of the first and second groups was noted. In all observations in animals of the first group, the periosteal cells were spindle-shaped, tightly adjacent to one another and located in parallel (Fig. 5a). In the second group, the proliferating cells of the periosteum had a stellate (processed) shape with clearly defined processes of the cytoplasm. Periosteal cells of animals of the second group, in contrast to the periosteal cells of animals of the first group, during the cultivation process formed a subconfluent, which had a “mesh” structure (see Fig. 5b).
When detecting ALP production by cultured periosteal cells, a positive reaction was observed only in animals of the first group (Fig. 6). Periosteal cells of animals of the second group did not produce ALP.


It is very significant that a similar picture was observed in the study of periosteal tissues obtained during surgical treatment of victims with osteoreparation problems after bone fractures. Thus, in the process of culturing periosteal tissues taken from patientsduring surgical treatment, the formation of a monolayer formed by heterogeneous cells was observed. In the monolayer, both elongated spindle-shaped cells and round-shaped and large cells with cytoplasmic projections were observed (Fig. 7). In its morphological picture, the monolayer formed by these cells resembled the monolayer formed by the cells of animals of the second group. The reaction with BCIP/NBT showed that among the proliferating cells there are single spindle-shaped cells that weakly produce alkaline phosphatase.

When studying the proliferation of endosteal cells, a picture similar to the proliferation of periosteal cells was observed. Endosteal cells began to proliferate on days 7–8 of cultivation. The number of proliferating endosteal cells, as in the case of periosteal cells, at different levels from the fracture line in the two groups was different (Table 3, Fig. 8).

In animals of the first group, the number of proliferating endosteal cells along the entire length of the bone fragment was approximately the same and ranged from 35 to 50 in the field of view. In animals of the second group, in the area of the fracture, the number of endosteal cells did not exceed 12-13 in the field of view, and in areas remote from the fracture it increased 3-4 times and ranged from 33 to 49 cells. In animals of this group, significant differences in the number of proliferating cells were noted only between levels 1-2 and level 3 (t=2.7, p0.02). There were no differences between levels 3, 4, and 5 (t=0.4, p> 0.10; t=0.23, p> 0.10, respectively). A comparison of the number of cells at levels 1–5 in animals of the two groups showed the presence of significant differences only at the first two levels - t=4.24, р<0.001 for the first level and t=2.74, р<0.02 for the second level. The morphology of endosteal cells in animals of the first and second groups was different. In animals of the first group, the cells had a round shape, characteristic of osteoblastic cells (Fig. 9a). In animals of the second group, the proliferating cells resembled fibroblast-like cells (Fig. 9b).

When culturing endosteal cells from animals of two groups, it was found that the proliferative activity of endosteal cells in animals of the two groups had significant differences. In animals of the first group, the proliferative activity of endosteal cells practically did not differ from control values (t = 2.7; p< 0.05). The proliferative activity of endosteal cells of animals of the second group was significantly higher than control values and the proliferative activity of endosteal cells of animals of the first group (t = 5.8; p< 0.001 and t = 5.4; p< 0.001, respectively) (Table 4).
By the 14th day of cultivation, the endosteal cells of the animals of the first group formed a subconfluent structure with osteoblast-like cells actively producing ALP (Fig. 10a). By this time, the endosteal cells of the animals of the second group had formed a dense monolayer formed by fibroblast-like cells that do not produce ALP (Fig. 10b).

Phase contrast microscopy of primary cultures of mesenchymal stromal cells isolated from the bone marrow canal of injured segments, it was noted that in animals of the first group, the number of MSCs adhering to the plastic was approximately 10-12 times greater than in animals of the second group (Fig. 11). Since only viable MSCs have the ability to adhere to plastic, microscopy data from primary cultures suggest that a significant number of MSCs die in the bone marrow canal under the influence of a high-intensity traumatic agent.

According to MTT analysis, the proliferative activity of MSCs on the 14th day of cultivation in animals of the first group was 3.5 times higher than control values (t= 9.9; p< 0.001), and in animals of the 2nd group – 2.5 times higher (t= 7.5; p< 0.001). The differences in the proliferative activity of MSCs between the groups were significant - the proliferative activity of MSCs from animals in the first group was 1.4 times higher than in animals from the second group (t = 3.2; p< 0.01) (Table 5).

By the 14th day of cultivation, MSCs from the CP of animals from the first group formed a dense monolayer with round-shaped cells, which in their morphology resembled epitheloid cells (Fig. 12a). By this time, MSCs from the CP of animals from the second group formed subconfluent. In its structure, the subconfluent formed by these cells resembled the subconfluent formed by the periosteal cells of animals of the same group (see Fig. 5b) - one part of the cells had a spindle-shaped shape, and the second part had a stellate shape (Fig. 12b).

When comparing the proliferative activity of various cellular sources of osteoreparation in animals of the control, first and second groups, it was noted that periosteal cells and MSCs, both in the control and in the first and second groups, have greater proliferative activity than endosteal cells. In animals of the control group, there were no differences between the proliferative activity of periosteal cells and MSCs (t = 0.63; p> 0.10), and in relation to the proliferative activity of endosteal cells, these differences were significant (t = 24.01; p< 0.001 and t = 27.69; p< 0.001, respectively) (Table 6, Fig. 13).

This difference is due to the fact that periosteal cells and MSCs are less differentiated than endosteal cells, which, in essence, are osteoblastic cells at different stages of differentiation - from preosteoblasts to osteoblasts.
After injury, the proliferative activity of cells from various sources of osteoreparation increased. The greatest activation of proliferation processes was observed in MSCs of the bone marrow canal. In animals of the first group, the proliferative activity of PC MSCs was 2.2 times higher than periosteal cells (t = 7.2; p< 0.001) and 11.3 times higher than endosteal cells (t = 12.55; p< 0.001). In animals of the second group, the proliferative activity of PC MSCs was 1.8 times greater than the proliferative activity of periosteal cells (t = 5.37; p< 0.001) and 4.3 times greater than that of endosteal cells (t = 9.49; p< 0.001). Periosteal cells were in second place in terms of proliferative activity. In the first group, their activity was 5.2 times higher than the activity of endosteal cells (t = 16.1; p< 0.001), and in the second group - 2.4 times higher (t = 12.7; p< 0.001). At the same time, the proliferative activity of periosteal cells and PC MSCs in animals of the first group was significantly higher than in animals of the second group (t = 2.19; p< 0.05 and t = 3.19; p< 0.01, respectively). On the part of endosteal cells, on the contrary, more pronounced proliferative activity was observed in animals of the second group (t = 5.35; р< 0.001).
The data obtained as a result of research indicate that the course of reparative processes in a bone wound is significantly influenced by the nature of the injury, namely, the intensity with which the traumatic agent affects the bone tissue. In low-intensity injuries, reparative processes developing in the bone wound lead to the formation of bone regenerate and fracture healing. With high-intensity injuries, reparative processes also occur in the bone wound, but they culminate in the formation of nonspecific fibrous tissue (see Fig. 2). The question naturally arises: “Why, after exposure to a high-intensity traumatic agent, does the formation of bone regenerate not occur in the fracture area?” In our opinion, the answer to this question is provided by the results of the research.
Data obtained from studies in in vitro conditions indicate that during high-intensity injuries, cells of the periosteum, endosteum and MSCs of the PC undergo certain morphofunctional changes. Firstly, with high-intensity trauma in bone fragments, a significant number of cellular elements of the periosteum and endosteum die at a considerable distance from the fracture line. The number of periosteal cells capable of proliferation in this area is reduced by more than 5 times, and the number of endosteal cells is reduced by more than 3.5 times. Most of the MSCs in the bone marrow canal also lose their viability. This is supported by a more than 10-fold decrease in the number of viable MSCs. Secondly, the proliferative activity of periosteal cells and MSCs during high-intensity injuries is 1.2 and 1.4 times lower than during low-intensity injuries, and the proliferative activity of endosteal cells, on the contrary, during high-intensity injuries is almost 2 times higher than during low-intensity injuries. The latter can be explained by the dedifferentiation of endosteal cells, which occurs as a result of exposure of bone tissue to a high-intensity traumatic agent. The dedifferentiation of endosteal cells is supported by a change in cell shape - from round, characteristic of osteoblastic cells, to spindle-shaped, characteristic of fibroblast-like cells, which have greater proliferative activity. Thirdly, with high-intensity injuries, periosteal cells and PC MSCs lose the ability to form a monolayer. Instead of monolayers formed by ordered cells of the same shape, subconfluent cultures are formed, formed by morphologically heterogeneous (including hypertrophied and necrotic) cells, which have a shape uncharacteristic for periosteal cells and MSCs (see Fig. 5, Fig. 7 and Fig. 12). Endosteal cells, unlike periosteal cells and MSCs, form a dense monolayer, but cells that have a fibroblast-like shape, which is not characteristic of endosteal cells, take part in the formation of the monolayer (see Fig. 10). In addition, as a result of exposure to a high-intensity traumatic agent, periosteal and endosteal cells stop producing ALP.
It is also important to note that cell cultures obtained from the group of animals with low-energy trauma are morphologically different from those obtained from animals with high-energy trauma. The former have epitheloid morphology (except for periosteal cells) and are osteoblastic lines (see Fig. 9a, 10a, 12a). Periosteal and endosteal cells of animals of the first group actively produce ALP, which indicates their osteogenic orientation and the ability to provide osteogenic induction of poorly differentiated bone marrow cells. At the same time, fibroblastoid cells of animals of the second group (see Fig. 9b, 10b, 12b) lose their osteogenic determination (as evidenced by their lack of ALP production) and the ability to provide osteogenic induction of poorly differentiated bone marrow cells.
In recent years, it has been shown that in a bone wound, under the influence of a significant amount of biologically active substances, activation and migration of proliferating non-committed poorly differentiated PC MSCs, which are essentially proto-myofibroblasts, occurs into the bone wound. In this case, a significant amount of TGF-β is locally released at the fracture site, a significant amount of the specific protein fibronectin accumulates, and powerful tensile forces of the extracellular matrix arise, reflecting the mechanical properties of the matrix and the remodeling activity of cells. Mechanoreception between myofibroblasts leads to significant contraction of granulation tissue formed in the wound. After this, a gradual reorganization of granulation tissue occurs with the launch of the process of apoptosis in myofibroblasts and the settlement of determined and induced cellular elements in their place.
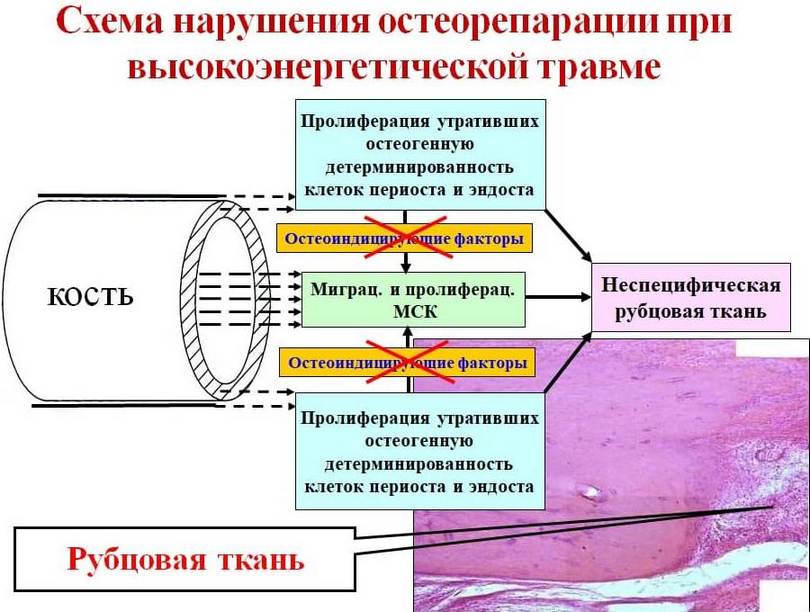
In fractures, the place of myofibroblasts that have undergone apoptosis is replaced by MSCs from the bone marrow canal and osteogenically determined elements of the endosteum and periosteum. Peristal and endosteal cells, along with the formation of bone regenerate along the periphery, osteoinduce MSCs that penetrate into the granulation tissue, thereby creating conditions for the formation of specific bone regenerate throughout the thickness of the bone wound (Fig. 12). In cases where, for some reason (in our case, as a result of exposure to high-intensity trauma), periosteal and endosteal cells lose their osteogenic determination, their osteoinducing effect on MSCs located in granulation tissue does not occur. As a result, during the reorganization of granulation tissue of a bone wound, in place of myofibroblasts that have undergone apoptosis, fibroblast-like cells migrate from the bone marrow canal, Haversian canals and the periosteum region, morphologically changed and having lost their inherent osteogenic properties, which contribute to the formation of a coarse fibrous nonspecific scar at the fracture site (Fig. 13).


Thus, summarizing all of the above, we can say that one of the main reasons that disrupts the course of reparative osteogenesis in high-intensity injuries is morphofunctional changes in the cellular sources of osteoreparation. The latter are characterized by dedifferentiation of osteogenically determined cells of the periosteum and endosteum, which leads to the development of osteogenic failure, which was described by V.G. Gololobov et al. based on empirical findings and confirmed by our experimental studies. The mechanisms leading to morphofunctional changes in cellular sources of osteoreparation are unknown to us and require separate study.
The problem of treating post-traumatic bone dysregeneration is the subject of a significant number of works, which consider both surgical and non-surgical methods of stimulating reparative osteogenesis. However, despite a fairly high percentage of successful treatment of patients with impaired consolidation of bone fragments, in 4.8 - 15.4% of patients it is not possible to achieve positive results. The greatest difficulties in the treatment of osteoreparation disorders arise after injuries resulting from exposure to a high-energy traumatic agent. According to various sources, in 10.8 - 13.7% of patients with high-energy injuries of the lower extremities, fractures did not heal within 1.5 - 3 years. It is known that the formation of callus in the fracture area occurs due to the progenitor cells of the periosteum, endosteum and mesenchymal stromal (stem) cells of the bone marrow cavity (MSCs). One of the determining mechanisms of callus formation is osteogenic differentiation of precursor cells proliferating into the bone wound. Recent studies have shown that differentiation of MSCs according to the osteoblastic type is a complex, multifaceted process and is carried out under the control of transcription factors, cellular and matrix interactions, systemic and local factors. The processes of osteogenic differentiation are significantly influenced by a large number of local factors, the vast majority of which are produced by cells of the osteoblastic line. But, despite the fact that in the process of osteogenic differentiation of precursor cells migrating into a bone wound, paracrine factors produced by periosteal and endosteal cells that already have an osteogenic orientation play a leading role, studies devoted to the study of changes in the morphofunctional state of cellular sources of osteoreparation depending on the mechanism of injury and the role of these changes in the development of dysregeneration could not be found in the available literature.
In this regard, a study was carried out of the role of changes in the morphofunctional state of cellular sources of osteoreparation (periosteum, endosteum and mesenchymal stromal cells of the bone marrow cavity (MCCs) in the disruption of reparative osteogenesis. For this purpose, experimental and clinical studies were carried out. The morphological picture of the bone wound area and the morphofunctional state of cellular sources of osteoreparation (periosteum, endosteum and MSCs) were studied KP).
Experimental studies were carried out in vivo and in vitro on 80 mature white outbred male rats. The animals were divided into three groups – control, first and second. Ten animals formed the control group. In these animals, the proliferative activity of periosteal, endosteal and PC MSCs was studied using MTT analysis. For this purpose, after removal from the experiment in intact animals, both legs were separated under aseptic conditions and the tibia bones were isolated from the soft tissues. The tibia bones were placed in a special preservative and kept in it for 24 hours. Then, under sterile conditions, the metaepiphyseal regions were cut off with scissors, leaving only the diaphyseal part of the bone for further examination. To determine the proliferative activity of PC MSCs under sterile conditions, the bone marrow contents were washed out of the bone marrow canal using an insulin syringe and physiological solution into Petri dishes with a culture medium. To simplify this procedure, the diaphyseal part of the bone was cut transversely in half. Half of the bone fragments were dissected longitudinally with ophthalmic scissors, thoroughly washed from the remaining bone marrow contents with physiological solution and placed in Petri dishes with a collagen substrate and nutrient medium so that only the periosteum was in contact with the collagen substrate (Fig. 1a). This position of bone fragments in Petri dishes created conditions for the proliferation of only periosteal cells. The remaining half of the bone fragments were carefully cleared from the periosteum using a scalpel, washed with saline and placed in Petri dishes with a collagen substrate and nutrient medium so that the bone tissue was in contact with the collagen substrate (Fig. 1 b). This created conditions for the proliferation of only cells of the bone tissue itself (endosteal cells).
Petri dishes with bone marrow contents and bone fragments were placed in a CO2-incubator and cultured under standard conditions (temperature 37°C and 5% CO2 content) with the nutrient medium changed every third day. After the start of cell proliferation in Petri dishes with bone fragments, the latter were carefully removed, and the cells attached to the collagen substrate continued to be cultured. MTT analysis of the proliferative activity of periosteal, endosteal and PC MSCs was carried out on the 14th day of cultivation. The results obtained were accepted as control ones.
Low- and high-energy fractures of the tibia were modeled in animals of the first (35 rats) and second (35 rats) groups. When simulating low-energy injuries (first group) under aseptic conditions, under ether anesthesia in the area of the middle third of the leg, a skin incision was made with a scalpel and the tibia was isolated. An osteotomy of the bone in the middle third was performed with a narrow chisel, after which the wound was sutured tightly. When simulating high-energy fractures (group 2), under ether anesthesia, a dosed blow was applied in the area of the middle third of the leg using a spring device, the force of which averaged 2.55±0.04 kg×m/s. In 10 animals (5 animals in each group), the injured segments were fixed paraosseously using needles for intramuscular injections and polyvinyl chloride tubes for intravenous infusions. These animals were removed from the experiment on the 7th day after injury. After the animals were removed from the experiment, the injured segment was separated under aseptic conditions, freed from the skin, fixed in a 10% solution of neutral formaldehyde, decalcified and stained with hematoxylin and eosin. The resulting histological preparations were studied under a light microscope. Sixty animals (30 animals from each group) were removed from the experiment 1 hour after injury. After the animals were removed from the experiment, the injured segment was separated under aseptic conditions, bone fragments were carefully isolated from the soft tissues and placed in a bottle with a sterile preservative. The bone fragments were kept in a bottle with a preservative for one day. Then, the same manipulations were performed with the bone fragments of each of the animals as in the animals of the control group and were cultured in a CO2 incubator under standard conditions. On days 5–8 of cultivation, periosteal and endosteal cells proliferating from bone fragments were counted using phase-contrast microscopy. Cells were counted in the field of view of the microscope at 100× magnification at five levels from the fracture line. The field of view of the microscope was used as a criterion to determine the level of counting. On the 14th day of cultivation in 45 Petri dishes of each group (15 dishes for each cell culture), the proliferative activity of periosteal, endosteal and PC MSCs was determined using MTT analysis. In the remaining 45 Petri dishes of each group (15 dishes for each cell culture), after the formation of a monolayer, the morphological state of proliferating cells of the periosteum, endosteum and PC MSCs was assessed using phase-contrast microscopy. In a monolayer of periosteal and endosteal cultures, the presence or absence of alkaline phosphatase (ALP) production by cells was determined using a reaction with BCIP/NBT Liquid Substrate System (Sigma, USA), which made it possible to judge the presence or absence of osteogenic determination (directionality) of cells.
Clinical studies were conducted in 5 patients with long-term nonunion of fractures of the long bones of the lower extremities. 2 patients had ununited fractures of the femur, and 3 patients had ununited fractures of the tibia. The duration of damage to the integrity of bone tissue in these patients ranged from 11 months to 2 years. During surgical treatment in patients, periosteal tissue measuring 0.5 x 0.5 cm was taken from the proximal bone fragment in the area of the fracture. The biopsy material was placed in a bottle with a preservative and kept for 24 hours. Then, under sterile conditions, sections of the periosteum were removed from the preservative and placed in Petri dishes with a nutrient medium and cultured in a CO2 incubator under standard conditions. After the appearance of cell proliferation, the periosteal tissues were removed, and the cells attached to the plastic were cultured until a monolayer was formed. Using phase-contrast microscopy, the morphological state of proliferating periosteal cells was assessed, and their osteogenic determination was assessed using the reaction to alkaline phosphatase.
Photo documentation of cell cultures and histo-preparations was carried out using a Leica DMIL inverted microscope, a Leica QWin500 Standart image processing workstation (version 2.3, sir. No. 3069) and a Sanyo TK-C1380 video camera (Germany). The digital material obtained as a result of the research was subjected to statistical processing.
When studying histomorphological preparations of the bone wound area on the 7th day after the injury, it was noted that the condition of the bone fragments and the healing of fractures in animals of the first and second groups was different. In animals of the first group, bone fragments had a significant number of cellular elements with well-stained nuclei. In the area of the bone wound, primitive bone beams formed, characteristic of the processes of callus formation (Fig. 2a). In the second group, the number of cellular elements in the bone tissue of the fragments was many times less than in the first group. They were located predominantly in areas of bone tissue remote from the fracture line. In the bone immediately adjacent to the fracture line, there were single cells with weakly stained nuclei. These changes morphologically resembled the picture of osteonecrosis. In the area of the fracture, reparative processes manifested themselves in the form of the formation of coarse-fibrous reticular connective tissue filling the bone wound (see Fig. 2b).
Thus, data from histomorphological studies indicate that the intensity of the traumatic agent affecting the bone affects the condition of the bone tissue and the course of fracture healing. With low-energy injuries in a bone wound, reparative processes occur with the formation of a specific bone regenerate, and with high-energy injuries, reparative processes are limited to the formation of nonspecific scar regenerate.
When culturing periosteum cells active cell proliferation began on the 5-6th day. In animals of the first group, periosteal cells proliferated evenly along the entire bone fragment (Fig. 3a). In animals of the second group, the proliferation of periosteal cells along the bone fragment was uneven. In the area adjacent to the fracture line, single cells proliferated. Active cell proliferation was observed in the area furthest from the fracture line (Fig. 3b).
The results of counting the number of periosteal cells in animals of groups 1 and 2 are presented in Table 1 and in the diagram (Fig. 4).
The presented data indicate that in animals of the first group the number of proliferating cells was the same throughout the entire bone fragment and amounted to 27-29 cells in the field of view. In the second group of animals, the number of proliferating cells in the fracture area did not exceed 6 in the field of view. In zones more distant from the fracture line, the number of proliferating cells gradually increased from 17 to 54 cells. The differences in the number of cells between levels from the fracture line in animals of the second group were significant (between levels 1, 2 and 3 - t=2.39, p<0.05; between levels 3 and 4 - t=2.3, p<0.05; between levels 4 and 5 - t=2.2, p<0.05).
A comparison of the number of cells at levels 1–5 in animals of the two groups showed that in animals of the first group, at the first three levels, the number of proliferating cells significantly exceeded that in animals of the second group (p< 0.002 andp < 0.05). At the fourth level, the number of proliferating periosteal cells in both groups was approximately the same (p> 0.10), and at the fifth level, the differences had an inverse relationship with the fracture line. In this area, the number of proliferating cells in the second group was significantly greater than in the first group (p<0.05).
During the cultivation process, the proliferative activity of periosteal cells in animals of the two groups was higher than the control value. The proliferative activity of periosteal cells in the first group was 1.6 times (t= 7.8; p< 0.001), and in the second group it was 1.4 times (t= 7.7; p< 0.001) higher than control values. The differences between the proliferative activity of periosteal cells of the first and second groups also had significant differences, although they were less pronounced (t = 2.2; p< 0.05) (Table 2).
During the formation of a cell monolayer, the presence of morphological differences between the cells of the periosteum in rats of the first and second groups was noted. In all observations in animals of the first group, the periosteal cells were spindle-shaped, tightly adjacent to one another and located in parallel (Fig. 5a). In the second group, the proliferating cells of the periosteum had a stellate (processed) shape with clearly defined processes of the cytoplasm. Periosteal cells of animals of the second group, in contrast to the periosteal cells of animals of the first group, during the cultivation process formed a subconfluent, which had a “mesh” structure (see Fig. 5b).
When detecting ALP production by cultured periosteal cells, a positive reaction was observed only in animals of the first group (Fig. 6). Periosteal cells of animals of the second group did not produce ALP.
It is very significant that a similar picture was observed in the study of periosteal tissues obtained during surgical treatment of victims with osteoreparation problems after bone fractures. Thus, in the process of culturing periosteal tissues taken from patientsduring surgical treatment, the formation of a monolayer formed by heterogeneous cells was observed. In the monolayer, both elongated spindle-shaped cells and round-shaped and large cells with cytoplasmic projections were observed (Fig. 7). In its morphological picture, the monolayer formed by these cells resembled the monolayer formed by the cells of animals of the second group. The reaction with BCIP/NBT showed that among the proliferating cells there are single spindle-shaped cells that weakly produce alkaline phosphatase.
When studying the proliferation of endosteal cells, a picture similar to the proliferation of periosteal cells was observed. Endosteal cells began to proliferate on days 7–8 of cultivation. The number of proliferating endosteal cells, as in the case of periosteal cells, at different levels from the fracture line in the two groups was different (Table 3, Fig. 8).
In animals of the first group, the number of proliferating endosteal cells along the entire length of the bone fragment was approximately the same and ranged from 35 to 50 in the field of view. In animals of the second group, in the area of the fracture, the number of endosteal cells did not exceed 12-13 in the field of view, and in areas remote from the fracture it increased 3-4 times and ranged from 33 to 49 cells. In animals of this group, significant differences in the number of proliferating cells were noted only between levels 1-2 and level 3 (t=2.7, p0.02). There were no differences between levels 3, 4, and 5 (t=0.4, p> 0.10; t=0.23, p> 0.10, respectively). A comparison of the number of cells at levels 1–5 in animals of the two groups showed the presence of significant differences only at the first two levels - t=4.24, р<0.001 for the first level and t=2.74, р<0.02 for the second level. The morphology of endosteal cells in animals of the first and second groups was different. In animals of the first group, the cells had a round shape, characteristic of osteoblastic cells (Fig. 9a). In animals of the second group, the proliferating cells resembled fibroblast-like cells (Fig. 9b).
When culturing endosteal cells from animals of two groups, it was found that the proliferative activity of endosteal cells in animals of the two groups had significant differences. In animals of the first group, the proliferative activity of endosteal cells practically did not differ from control values (t = 2.7; p< 0.05). The proliferative activity of endosteal cells of animals of the second group was significantly higher than control values and the proliferative activity of endosteal cells of animals of the first group (t = 5.8; p< 0.001 and t = 5.4; p< 0.001, respectively) (Table 4).
By the 14th day of cultivation, the endosteal cells of the animals of the first group formed a subconfluent structure with osteoblast-like cells actively producing ALP (Fig. 10a). By this time, the endosteal cells of the animals of the second group had formed a dense monolayer formed by fibroblast-like cells that do not produce ALP (Fig. 10b).
Phase contrast microscopy of primary cultures of mesenchymal stromal cells isolated from the bone marrow canal of injured segments, it was noted that in animals of the first group, the number of MSCs adhering to the plastic was approximately 10-12 times greater than in animals of the second group (Fig. 11). Since only viable MSCs have the ability to adhere to plastic, microscopy data from primary cultures suggest that a significant number of MSCs die in the bone marrow canal under the influence of a high-intensity traumatic agent.
According to MTT analysis, the proliferative activity of MSCs on the 14th day of cultivation in animals of the first group was 3.5 times higher than control values (t= 9.9; p< 0.001), and in animals of the 2nd group – 2.5 times higher (t= 7.5; p< 0.001). The differences in the proliferative activity of MSCs between the groups were significant - the proliferative activity of MSCs from animals in the first group was 1.4 times higher than in animals from the second group (t = 3.2; p< 0.01) (Table 5).
By the 14th day of cultivation, MSCs from the CP of animals from the first group formed a dense monolayer with round-shaped cells, which in their morphology resembled epitheloid cells (Fig. 12a). By this time, MSCs from the CP of animals from the second group formed subconfluent. In its structure, the subconfluent formed by these cells resembled the subconfluent formed by the periosteal cells of animals of the same group (see Fig. 5b) - one part of the cells had a spindle-shaped shape, and the second part had a stellate shape (Fig. 12b).
When comparing the proliferative activity of various cellular sources of osteoreparation in animals of the control, first and second groups, it was noted that periosteal cells and MSCs, both in the control and in the first and second groups, have greater proliferative activity than endosteal cells. In animals of the control group, there were no differences between the proliferative activity of periosteal cells and MSCs (t = 0.63; p> 0.10), and in relation to the proliferative activity of endosteal cells, these differences were significant (t = 24.01; p< 0.001 and t = 27.69; p< 0.001, respectively) (Table 6, Fig. 13).
This difference is due to the fact that periosteal cells and MSCs are less differentiated than endosteal cells, which, in essence, are osteoblastic cells at different stages of differentiation - from preosteoblasts to osteoblasts.
After injury, the proliferative activity of cells from various sources of osteoreparation increased. The greatest activation of proliferation processes was observed in MSCs of the bone marrow canal. In animals of the first group, the proliferative activity of PC MSCs was 2.2 times higher than periosteal cells (t = 7.2; p< 0.001) and 11.3 times higher than endosteal cells (t = 12.55; p< 0.001). In animals of the second group, the proliferative activity of PC MSCs was 1.8 times greater than the proliferative activity of periosteal cells (t = 5.37; p< 0.001) and 4.3 times greater than that of endosteal cells (t = 9.49; p< 0.001). Periosteal cells were in second place in terms of proliferative activity. In the first group, their activity was 5.2 times higher than the activity of endosteal cells (t = 16.1; p< 0.001), and in the second group - 2.4 times higher (t = 12.7; p< 0.001). At the same time, the proliferative activity of periosteal cells and PC MSCs in animals of the first group was significantly higher than in animals of the second group (t = 2.19; p< 0.05 and t = 3.19; p< 0.01, respectively). On the part of endosteal cells, on the contrary, more pronounced proliferative activity was observed in animals of the second group (t = 5.35; р< 0.001).
The data obtained as a result of research indicate that the course of reparative processes in a bone wound is significantly influenced by the nature of the injury, namely, the intensity with which the traumatic agent affects the bone tissue. In low-intensity injuries, reparative processes developing in the bone wound lead to the formation of bone regenerate and fracture healing. With high-intensity injuries, reparative processes also occur in the bone wound, but they culminate in the formation of nonspecific fibrous tissue (see Fig. 2). The question naturally arises: “Why, after exposure to a high-intensity traumatic agent, does the formation of bone regenerate not occur in the fracture area?” In our opinion, the answer to this question is provided by the results of the research.
Data obtained from studies in in vitro conditions indicate that during high-intensity injuries, cells of the periosteum, endosteum and MSCs of the PC undergo certain morphofunctional changes. Firstly, with high-intensity trauma in bone fragments, a significant number of cellular elements of the periosteum and endosteum die at a considerable distance from the fracture line. The number of periosteal cells capable of proliferation in this area is reduced by more than 5 times, and the number of endosteal cells is reduced by more than 3.5 times. Most of the MSCs in the bone marrow canal also lose their viability. This is supported by a more than 10-fold decrease in the number of viable MSCs. Secondly, the proliferative activity of periosteal cells and MSCs during high-intensity injuries is 1.2 and 1.4 times lower than during low-intensity injuries, and the proliferative activity of endosteal cells, on the contrary, during high-intensity injuries is almost 2 times higher than during low-intensity injuries. The latter can be explained by the dedifferentiation of endosteal cells, which occurs as a result of exposure of bone tissue to a high-intensity traumatic agent. The dedifferentiation of endosteal cells is supported by a change in cell shape - from round, characteristic of osteoblastic cells, to spindle-shaped, characteristic of fibroblast-like cells, which have greater proliferative activity. Thirdly, with high-intensity injuries, periosteal cells and PC MSCs lose the ability to form a monolayer. Instead of monolayers formed by ordered cells of the same shape, subconfluent cultures are formed, formed by morphologically heterogeneous (including hypertrophied and necrotic) cells, which have a shape uncharacteristic for periosteal cells and MSCs (see Fig. 5, Fig. 7 and Fig. 12). Endosteal cells, unlike periosteal cells and MSCs, form a dense monolayer, but cells that have a fibroblast-like shape, which is not characteristic of endosteal cells, take part in the formation of the monolayer (see Fig. 10). In addition, as a result of exposure to a high-intensity traumatic agent, periosteal and endosteal cells stop producing ALP.
It is also important to note that cell cultures obtained from the group of animals with low-energy trauma are morphologically different from those obtained from animals with high-energy trauma. The former have epitheloid morphology (except for periosteal cells) and are osteoblastic lines (see Fig. 9a, 10a, 12a). Periosteal and endosteal cells of animals of the first group actively produce ALP, which indicates their osteogenic orientation and the ability to provide osteogenic induction of poorly differentiated bone marrow cells. At the same time, fibroblastoid cells of animals of the second group (see Fig. 9b, 10b, 12b) lose their osteogenic determination (as evidenced by their lack of ALP production) and the ability to provide osteogenic induction of poorly differentiated bone marrow cells.
In recent years, it has been shown that in a bone wound, under the influence of a significant amount of biologically active substances, activation and migration of proliferating non-committed poorly differentiated PC MSCs, which are essentially proto-myofibroblasts, occurs into the bone wound. In this case, a significant amount of TGF-β is locally released at the fracture site, a significant amount of the specific protein fibronectin accumulates, and powerful tensile forces of the extracellular matrix arise, reflecting the mechanical properties of the matrix and the remodeling activity of cells. Mechanoreception between myofibroblasts leads to significant contraction of granulation tissue formed in the wound. After this, a gradual reorganization of granulation tissue occurs with the launch of the process of apoptosis in myofibroblasts and the settlement of determined and induced cellular elements in their place.
In fractures, the place of myofibroblasts that have undergone apoptosis is replaced by MSCs from the bone marrow canal and osteogenically determined elements of the endosteum and periosteum. Peristal and endosteal cells, along with the formation of bone regenerate along the periphery, osteoinduce MSCs that penetrate into the granulation tissue, thereby creating conditions for the formation of specific bone regenerate throughout the thickness of the bone wound (Fig. 12). In cases where, for some reason (in our case, as a result of exposure to high-intensity trauma), periosteal and endosteal cells lose their osteogenic determination, their osteoinducing effect on MSCs located in granulation tissue does not occur. As a result, during the reorganization of granulation tissue of a bone wound, in place of myofibroblasts that have undergone apoptosis, fibroblast-like cells migrate from the bone marrow canal, Haversian canals and the periosteum region, morphologically changed and having lost their inherent osteogenic properties, which contribute to the formation of a coarse fibrous nonspecific scar at the fracture site (Fig. 13).
Thus, summarizing all of the above, we can say that one of the main reasons that disrupts the course of reparative osteogenesis in high-intensity injuries is morphofunctional changes in the cellular sources of osteoreparation. The latter are characterized by dedifferentiation of osteogenically determined cells of the periosteum and endosteum, which leads to the development of osteogenic failure, which was described by V.G. Gololobov et al. based on empirical findings and confirmed by our experimental studies. The mechanisms leading to morphofunctional changes in cellular sources of osteoreparation are unknown to us and require separate study.